Review Article | Volume 1 - Issue 2 | Article DOI :
Download PDF
Liujun Chen1 , Shanshan Xu1 , Luoshiyuan Zuo3 , Song Han1,2, Jun Yin1,2, Biwen Peng1,2, Xiaohua He1,2 and Wanhong Liu1,2*
1Hubei Province Key Laboratory of Allergy and Immunology, School of Basic Medical Sciences, Wuhan University, China
2Hubei Provincial Key Laboratory of Developmentally Originated Disease, China
3School of Health Sciences, Wuhan University, China
Corresponding Author:
Wanhong Liu, School of Basic MedicalSciences, Wuhan University, China,Tel: 0086-27-68759985,Fax: 0086-27-68759991,
Keywords
HIV; Long noncoding RNAs (lncRNAs); Function; Latency; Antivirus
Abstract
The Human Immunodeficiency Virus (HIV) is a retrovirus that has been aroused worldwide concern, due to its chronic and persistent infection, ultimately leading to a causal result of Acquired Immunodeficiency Syndrome (AIDS). Long noncoding RNAs (lncRNAs) are non-protein coding transcript longer than 200 Base Pairs (bp) and being considered to be key regulators that involved in various biological processes, such as chromatin modification, transcriptional regulation, post-transcriptional regulation, intracellular trafficking and etc. What deserves to be noticed is that lncRNAs are recently being reported to link with viruses closely, and lncRNAs are differentially expressed after a variety of virus infections, including HIV-1 infection. In this paper, we review the rapidly advancing field of lncRNAs, focus on the current progress of lncRNAs in HIV-1 infection, and briefly discuss their different roles in host gene regulation and viral replication during the establishment or maintenance of viral latency. Interestingly, lncRNAs may emerge as novel biomarkers of antiviral drugs and provide potential targets for new therapeutics of AIDS.
Citation
Chen L, Xu S, Zuo L, Han S, Yinc J, Peng B, et al. LncRNA: A Rising Star in Virus-Host Cross-Talk during HIV-1 Infection. SM Virol. 2016; 1(2): 1010.
Keywords
HIV; Long noncoding RNAs (lncRNAs); Function; Latency; Antivirus
Introduction
The Human Immunodeficiency Virus (HIV) is a RNA virus belonging to the genera of lentiviruses, family of retrovirus that is characterized by its chronic and persistent infection. HIV ultimately leads to a causal result of Acquired Immunodeficiency Syndrome (AIDS), which persists as one of the greatest global health challenge [1,2], even without an efficacious vaccine in the near future. It is especially imminent and vital to explore and improve the methods of searching novel host targets and biomarkers for AIDS prognostics and diagnostics. In recent years, different types of noncoding RNAs (ncRNAs), including lncRNAs have been discovered that involved in various biological processes, including HIV infection [3-6]. In this paper, we focus on these phenomena, compared these IncRNAs and illustrated the intricate relationship between IncRNAs and HIV.
LncRNA
LncRNA synopsis
It is worth mentioning that there are a growing number of reports describing long noncoding RNA, although the current literatures are dominated by short transcripts. Known as a particular noncoding transcriptional class without any single unifying feature, lncRNAs are currently referred as RNA transcripts of longer than 200 Base Pairs (bp), without evident protein-coding capability. To the extent that lncRNA was initially regarded as ‘junk’ RNA or spurious transcriptional noise in the last few years. But this view was based on misconceptions and had been outdated stereotypes [7-10].
LncRNAs have a broad definition encompassing different classes of RNA transcripts [9,11]. Based on transcript length, a threshold of 200 base pairs separates long from short ncRNAs, which is the most commonly used attribute for their classification [12,13]. Classification based on association with annotated serves as the foundation of the GENCODE classification of lncRNAs that are classified as intronic ncRNA, sense ncRNA, natural antisense ncRNA and etc [14-16]. Classification based on association with other DNA elements of known function, such as enhancer-associated RNA, promoter-associated long RNA, telomeric repeat-containing RNA and etc [12,17,18]. Classification based on sequence and structure conservation, such as transcribed-ultraconserved regions, Hypoxia-induced noncoding ultraconserved transcript, long-intergenic noncoding RNA, long-intervening noncoding RNA and etc [19-21].
LncRNAs are transcribed and processed in a manner similar to Messenger RNAs (mRNAs) that have been demonstrated to be important in development and disease. Additionally, lncRNAs involved in a broad spectrum of gene regulatory functions such as epigenetic activities, translational regulation or scaffolding/decoy functions that have been widely acknowledgement.[22,23]. Notablely, lncRNAs are reported to be closely related to virus infections.
As lncRNA being a ring star of important molecular, plentiful researches and facts make more researchers realize lncRNA may have functions in virus infection and host defense [24-28]. For example, after influenza-A virus and Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) infections, there were 5,329 IncRNAs expressing differentially [24]. In Coxsackievirus A16 (CA16) infected Rhabdomyosarcoma (RD) cells identified by RNA-seq, there were 1,970 lncRNAs were found to have differential expression [25]. Besides, more than 4,800 lncRNAs were observed the differential expression in Enterovirus 71 (EV71) infections [26]. Conversely, lncRNA GAS5 can inhibit Hepatitis C Virus (HCV) replication by binding viral NS3 protein [27]. Theiler’s Murine Encephalomyelitis Virus (TMEV) infection can be influence by lncRNA NeST susceptibility [28].
LncRNAs changed by HIV infection
Two separate researches were choreographed by two independent groups that utilized the next generation sequencing to reveal a map of lncRNAs information in different cells after HIV infection [3,4]. Plenty of lncRNAs were identified to have significant differential expression in different cells with HIV-1 infected by qRT-PCR. Zhang et al. identified that 8 lncRNAs were upregulated and 5 lncRNAs were downregulated in HIV-1-infected Jurkat cells, while 6 lncRNAs were upregulated and 5 lncRNAs were downregulated in HIV-1-infected MT4 cells. In contrast, Imam et al. found that 2 lncRNAs were downregulated and 21 lncRNAs were upregulated in HIV-1-infected Jurkat cells. In addition, 4 lncRNAs were downregulated and 23 lncRNAs were upregulated in the PMA-activated J1.1 cells compared to mock-activated cells.
Intriguingly, differential expressed lncRNA SRA was reported in these two groups. Zhang et al. found that lncRNA SRA was downregulated in both Jurkat and MT4 cells [3]. However, Imam et al. found that lncRNA SRA was upregulated in both Jurkat and J1.1 cells [4]. Such differences may imply that lncRNA differential expression vary from cell to cell at different post-infection time. These data suggested that the above lncRNAs may be modulated by viruses itself or cellular protein during HIV infection process. But more work is needed to study on their regulatory mechanism.
p53 dependent apoptosis-related lncRNAs in HIV
As is mentioned above, a map of lncRNAs information, revealed by two unique researches, also identified that two p53-dependent lncRNAs (lincRNA-p21 and PANDA) were significantly differentially expressed after HIV infection. PANDA was downregulation [3], while lincRNA-p21 was upregulation [4]. PANDA is an antisense lncRNA co-regulated together with CDKN1A by tumor suppressor protein p53, a promoter of CDKN1A, related to DNA damage activated [29]. Overexpression of PANDA inhibits apoptosis by binding to NF-YA and preventing NF-YA co-localization at promoter regions of apoptotic genes, suggesting that DNA damage activates p53-mediated transcription at CDKN1A and PANDA that functions synergistically to mediate cell cycle arrest and survival [29, 30]. LincRNA-p21 is a p53-induced large intergenic noncoding RNA, influences the p53 tumor suppressor pathway by acting in cis as a locus-restricted co-activator for p53-mediated p21 expression. Knockdown of lincRNA-p21 reduces the effect of apoptotic by p53 [31,32]. Whereas, recent researches suggests that HIV succeeds in inhibiting lincRNA-p21 and the p53 induced apoptotic pathway by activating the MAP2K1/ERK2 pathway, so as to better promote viral replication [33]. Thus, the results collectively suggest that p53 dependent apoptosis-related lncRNAs shows great relevance in HIV infection. These two effects were both closely related to promotion of apoptosis. Nevertheless, HIV replication requires the avoidance of the p53-apoptotic pathway, so there may be other kinds of pathway where HIV can suppress the p53 response.
Multifunctional role of lncRNA in HIV
infection (Table 1) Epigenetic modulators
Table 1: LncRNA functions in HIV-1 infection.
| Functions |
Name |
Target |
mechanism |
References |
| Epigenetic modulators |
HIV-derived antisense |
DNMT3a, |
Form H3K9me2, H3K27me3, histone deacetylation and other heterochromatin; |
[35] |
| lncRNA |
HIV 5’LTR |
Suppress the activity of viral promoter and gene. |
| |
NEAT1 |
paraspeckle |
Involve in HIV Rev-dependent nuclear export. |
[3] |
| Scaffolding/decoy regulator |
7SK RNA |
7SK RNP, |
Repress the kinase activity of CDK9; preventing recruitment of P-TEFb to the |
[31-41] |
| |
P-TEFb |
HIV-1 promoter. |
| |
NRON |
NFAT, Tat |
Regulate NFAT activity by changing its localization, prevent it binding to the |
[4,44,45] |
| |
viral 5’LTR; Links Tat to the ubiquitin/ proteasome for degradation. |
Much of the research in lncRNAs has indicated that viruses can modulate expression levels of host-derived lncRNAs during viral infection for host or viral gene regulation [3,4,25]. Conversely, viruses can also take advantage of their encoded lncRNAs to maintain latent or replication with complex regulatory mechanism [34,35]. An HIV-encoded antisense long non-coding RNA has been reported that epigenetically regulates viral transcription [35].
This lncRNA is located in the 5’LTR of HIV, which is the viral core promoter region, and interacted directly with DNA methyltransferase 3A (DNMT3a). Besides that, HIV-encoded antisense lncRNA recruit Histone Deacetylase(HDAC1), Histone Methyltransferases(EZH2) to viral 5’LTR to regulate the epigenetic state at the promoter region that form H3K9me2, H3K27me3, histone deacetylation and other heterochromatin thereby suppressing the activity of promoter and silencing subsequent viral gene transcription. Inhibition of this antisense lncRNA with small single stranded antisense RNAs (asRNAs) resulted in the activation of viral gene expression [35]. Thus, HIV-encoded antisense lncRNA modulate HIV gene expression by alternating the epigenetic landscape at the viral promoter that may contribute to latent infection.
Scaffolding/decoy regulator
NEAT1: NEAT1 RNA, whose full name is “nuclear paraspeckle assembly transcript 1”, is up-regulated following infection by HIV-1 [3], which is crucial to form nuclear structures termed paraspeckles [36]. The depletion of NEAT1 resulted in an increase in HIV-1 expression [3], and also led to obvious reduction in the number of paraspeckle bodies. In fact, NEAT1 has no effect on formation of RNA-binding proteins in the paraspeckle bodies, but change the distribution of core proteins, and thus influence the paraspeckles formation[37], suggesting that NEAT1 may act as a scaffolding molecule for recruiting nuclear proteins (p54nrb, PSF, and Matrin3, etc.) and enables the formation of the paraspeckles [38]. These three nuclear proteins (p54nrb, PSF, Matrin3) had been demonstrated to be involved in HIV Rev-dependent nuclear export of incompletely spliced viral RNAs containing cis-acting instability elements (INS) at the posttranscriptional level [3]. In other words, knockdown of NEAT1 increase nuclear cytoplasmic export and promote viral infections.
7SK RNA: 7SK RNA is an evolutionarily conserved small nuclear RNA (snRNA) with 331 bp [39]. As the definitions of length, 7SK RNA can be also defined as lncRNA. The same as a binding scaffold role, 7SK RNA was found to be associated with Cyclin T1, CDK9, HEXIM1, MEPCE, and LARP7 and formed 7SK RNP complexes [39-42]. In addition, as a specific Positive Transcription Elongation Factor B (P-TEFb) associated factor [40], 7SK RNA involved in HIV replication cycle by repressing the kinase activity of CDK9 [39], preventing recruitment of P-TEFb to the HIV-1 promoter [41]. Conversely, it doesn’t mean that the virus ignore the existence of this effect. The feedback is that HIV protein that not only attenuate the formation of 7SK RNP, but also has feature extraction function of Cyclin T1 and CDK9, consequently, activating the transcription of HIV proviral virus [41]. T-cell receptor signaling activates P-TEFb through ERK pathway or the PI3K/Akt pathway that would enhances Tat-dependent transcriptional elongation from latent HIV proviruses by releasing Cyclin T1/CDK9 from the 7SK RNP [42]. Hence, 7SK RNA may participate in establishment and maintenance of HIV latent infection by resting or activating CD4+ T cells through regulating 7SK RNP expression levels.
NRON: NRON, a long noncoding RNA with 2730 bp, known as a noncoding repressor of the Nuclear Factor of Activated T Cells (NFAT) [43], is significantly differential expression during HIV 1 infection that can participate in regulation of various physiologic functions in HIV-1 life cycle. NRON is decreased by the early viral accessory protein Nef and intriguingly increased by the HIV late protein Vpu [4].
Acting as a decoy role, NRON binds to multiple proteins, including Calmodulin-Binding Protein (IQGAP1), nuclear trans port factor (KPNB1), phosphatase (PPP2R1A), and the proteasome (PSMD11). NRON regulates NFAT activity by sequestering these aforementioned proteins, changing the subcellular localization of transcription factors NFAT, but not affect the transcription of the NFAT. NFAT can Promotes HIV transcriptional activity through its binding to the viral 5’LTR region. Depletion of NRON enhanced HIV-1 replication through increasing activity of NFAT and the viral LTR [44].
Furthermore, the latest research in the first half of 2016 indicated that NRON can also potently repress the viral transcription by decreasing the viral transactivator protein Tat. NRON directly links Tat to the ubiquitin/proteasome components, including cullin 4B (CUL4B) and PSMD11, thus accelerating Tat degradation. Knockdown of NRON, especially in combination with a HDAC inhibitor, remarkably reactivates the viral production from the HIV 1-latently infected primary CD4+ T lymphocytes [45]. Therefore, NRON may play a important role in HIV-1 latency maintenance as a decoy.
Perspective
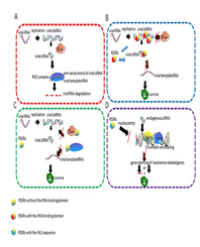
LncRNA may play a crucial role in regulating HIV infection. Here, we focused our discussion on 6 lncRNAs in HIV infection that has given researchers some latest molecular insight into how lncRNAs affect key regulatory pathways in HIV (Figure 1).

Figure 1: Model for role of lncRNA in HIV infection. (a) LncRNA can work as an epigenetic modulator by recruiting chromatin modifying complexes to specific genomic loci to impart their catalytic activity. In this case, HIV-derived antisense lncRNA interacts directly with DNA methyltransferase 3A (DNMT3a) and recruits histone deacetylase (HDAC1), histone methyltransferases (EZH2) to viral 5’LTR to form H3K9me2, H3K27me3, and histone deacetylation, induce heterochromatin and repress the activity of promoter, then silence subsequent viral gene transcription [35]. (b) LncRNA can regulate the transcriptional process through acting as a scaffolding/decoy regulator. In that case, (1) LncRNA NEAT1 recruits nuclear proteins (p54nrb, PSF, and Matrin3, etc.) to form paraspeckles, and avoids these protein involving in HIV Rev-dependent nuclear export [3]. (2) LncRNA 7SK recruits Cyclin T1, CDK9, HEXIM1, MEPCE, and LARP7 to form 7SK RNP complexes, repressing the kinase activity of CDK9 [37], preventing recruitment of P-TEFb to the HIV-1 promoter [39-41] (3) LncRNA NRON regulates NFAT activity by working as a decoy that change the subcellular localization calmodulin-binding protein (IQGAP1), nuclear trans-port factor (KPNB1), phosphatase (PPP2R1A) and proteasome (PSMD11), the key factors of NFAT, avoiding NFAT promoting viral transcriptional activity. Additionally, NRON can link Tat to the ubiquitin/proteasome components cullin 4B (CUL4B) and PSMD11 to accelerate Tat degradation [4,44,45].
With the development of research, a growing number of differential expressed lncRNAs would be found that involved in the process of virus infection, regulation of the host immune response to the virus, and cell growth process. Furthermore, all of these lncRNAs can be served as potential targets for novel antiviral drugs [46]. Based on the research of lncRNAs, a novel perspective of the virus-host interactions will come into view and may bring some new revelations to AIDS gene therapy.
Acknowledgments
This work was supported by the National Natural Sciences Foundation of China (No. 81371790, 81641093, 81371422, 81571481 and 31170154), Major AIDS and Viral Hepatitis and Other Major Infectious Disease Prevention and Control project of China (2014ZX10001003), the Fundamental Research Funds for the Central Universities of China and the Translational Medical Research Fund of Wuhan University School of Medicine.
References
1. Barre-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science. 1983; 220: 868-871.
2. Palella FJ, Jr., Delaney KM, Moorman AC, Loveless MO, Fuhrer J, Satten GA, et al. Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV Outpatient Study Investigators. The New England journal of medicine. 1998; 338: 853-860.
3. Zhang Q, Chen CY, Yedavalli VS, Jeang KT. NEAT1 long noncoding RNA and paraspeckle bodies modulate HIV-1 posttranscriptional expression. MBio. 2013; 4: e00596-00512.
4. Imam H, Bano AS, Patel P, Holla P, Jameel S. The lncRNA NRON modulates HIV-1 replication in a NFAT-dependent manner and is differentially regulated by early and late viral proteins. Scientific reports. 2015; 5: 8639.
5. Cheng Q, Dong L, Zhang F, Yuan P, Li Z, Sun Y, Yin J, et al. Short communication: Efficiently inhibiting HIV-1 replication by a prototype foamy virus vector expressing novel H1 promoter-driven short hairpin RNAs. AIDS research and human retroviruses. 2015; 31: 183-188.
6. Hou P, Chen S, Wang S, Yu X, Chen, Jiang M, Zhuang K, et al. Genome editing of CXCR4 by CRISPR/cas9 confers cells resistant to HIV-1 infection. Scientific reports. 2015; 5: 15577.
7. Johnsson P, Ackley A, Vidarsdottir L, Lui WO, Corcoran M, et al. A pseudogene long-noncoding-RNA network regulates PTEN transcription and translation in human cells. Nature structural & molecular biology. 2013; 20: 440-446.
8. Derrien T, Johnson R, Bussotti G, Tanzer A, Djebali S, Tilgner H, et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome research. 2012; 22: 1775 1789.
9. St Laurent G, Wahlestedt C, Kapranov P. The Landscape of long noncoding RNA classification. Trends in genetics: TIG. 2015; 31: 239-251.
10. Wang Z, Zhang XJ, Ji YX, Zhang P, Deng KQ, Gong J, et al. The long noncoding RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy. Nature medicine. 2016; 22: 1131-1139.
11. Cao J. the functional role of long non-coding RNAs and epigenetics. Biological procedures online. 2014; 16: 11.
12. Kapranov P, Cheng J, Dike S, Nix DA, Duttagupta R, Willingham AT, Stadler PF, et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science. 2007; 316: 1484-1488.
13. Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Molecular cell. 2011; 43: 904-914.
14. Kapranov P, Drenkow J, Cheng J, Long J, Helt G, et al. Examples of the complex architecture of the human transcriptome revealed by RACE and high-density tiling arrays. Genome research. 2005; 15: 987-997.
15. Nakaya HI, Amaral PP, Louro R, Lopes A, Fachel AA, Moreira YB, et al. Genome mapping and expression analyses of human intronic noncoding RNAs reveal tissue-specific patterns and enrichment in genes related to regulation of transcription. Genome Biol. 2007; 8: R43.
16. Katayama S, Tomaru Y, Kasukawa T, Waki K, Nakanishi M, et al. Antisense transcription in the mammalian transcriptome. Science. 2005; 309: 1564 1566.
17. Kim TK, Hemberg M, Gray JM, Costa AM, Bear DM, et al. Widespread transcription at neuronal activity-regulated enhancers. Nature. 2010; 465: 182-187.
18. Luke B, Lingner J. TERRA: telomeric repeat-containing RNA. EMBO journal. 2009; 28: 2503-2510.
19. Hudson RS, Yi M, Volfovsky N, Prueitt RL, Esposito D, Volinia S, et al. Transcription signatures encoded by ultraconserved genomic regions in human prostate cancer. Molecular cancer. 2013; 12: 13.
20. Ferdin J, Nishida N, Wu X, Nicoloso MS, Shah MY, et al. HINCUTs in cancer: hypoxia-induced noncoding ultraconserved transcripts. Cell death differs. 2013; 20: 1675-1687.
21. Guttman M, Amit I, Garber M, French C, Lin MF, David Feldser, et al. Chromatin signature reveals over a thousand highly conserved large non coding RNAs in mammals. Nature. 2009; 458: 223-227.
22. Garitano-Trojaola A, Agirre X, Prosper F, Fortes P. Long non-coding RNAs in haematological malignancies. International journal of molecular sciences. 2013; 14: 15386-15422.
23. Iyer MK, Niknafs YS, Malik R, Singhal U, Sahu A, Hosono Y, et al. The landscape of long noncoding RNAs in the human transcriptome. Nature genetics. 2015; 47: 199-208.
24. Josset L, Tchitchek N, Gralinski LE, Ferris MT, Eisfeld AJ, Green RR, et al. Annotation of long non-coding RNAs expressed in collaborative cross founder mice in response to respiratory virus infection reveals a new class of interferon-stimulated transcripts. RNA biology. 2014; 11: 875-890.
25. Shi Y, Tu H, Chen X, Zhang Y, Chen L, Liu Z, Sheng J, et al. The long non-coding RNA expression profile of Coxsackievirus A16 infected RD cells identified by RNA-seq. Virologica Sinica. 2016; 31: 131-141.
26. Yin Z, Guan D, Fan Q, Su J, Zheng W, et al. lncRNA expression signatures in response to enterovirus 71 infection. Biochemical and biophysical research communications. 2013; 430: 629-633.
27. Qian X, Xu C, Zhao P, Qi Z. Long non-coding RNA GAS5 inhibited hepatitis C virus replication by binding viral NS3 protein. Virology. 2016; 492: 155-165.
28. Gomez JA, Wapinski OL, Yang YW, Bureau JF, Gopinath S, Monack DM, et al. The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-gamma locus. Cell. 2013; 152: 743-754.
29. Hung T, Wang Y, Lin MF, Koegel AK, Kotake Y, et al. Extensive and coordinated transcription of noncoding RNAs within cell-cycle promoters. Nature genetics. 2011; 43: 621-629.
30. Puvvula PK, Desetty RD, Pineau P, Marchio A, Moon A, et al. Long noncoding RNA PANDA and scaffold-attachment-factor SAFA control senescence entry and exit. Nature communications. 2014; 5: 5323.
31. Huarte M, Guttman M, Feldser D, Garber M, Koziol MJ, Kenzelmann-Broz D, et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell. 2010; 142: 409-419.
32. Bao X, Wu H, Zhu X, Guo X, Hutchins AP, Luo Z, et al. The p53-induced lincRNA-p21 derails somatic cell reprogramming by sustaining H3K9me3 and CpG methylation at pluripotency gene promoters. Cell research. 2015; 25: 80-92.
33. Barichievy S, Naidoo J, Mhlanga MM. Non-coding RNAs and HIV: viral manipulation of host dark matter to shape the cellular environment. Frontiers in genetics. 2015; 6: 108.
34. Kobayashi-Ishihara M, Yamagishi M, Hara T, Matsuda Y, Takahashi R, et al. HIV-1-encoded antisense RNA suppresses viral replication for a prolonged period. Retrovirology. 2012; 9: 38.
35. Saayman S, Ackley A, Turner AM, Famiglietti M, Bosque A, et al. An HIV encoded antisense long noncoding RNA epigenetically regulates viral transcription. Molecular therapy : the journal of the American Society of Gene Therapy. 2014; 22: 1164-1175.
36. Sunwoo JS, Lee ST, Im W, Lee M, Byun JI, Jung KH, et al. Altered Expression of the Long Noncoding RNA NEAT1 in Huntington’s Disease. Mol neurobiology. 2016.
37. Choudhry H, Albukhari A, Morotti M, Haider S, Moralli D et al. Tumor hypoxia induces nuclear paraspeckle formation through HIF-2alpha dependent transcriptional activation of NEAT1 leading to cancer cell survival. Oncogene. 2015; 34: 4546.
38. Clemson CM, Hutchinson JN, Sara SA, Ensminger AW, Fox AH, et al. An architectural role for a nuclear noncoding RNA: NEAT1 RNA is essential for the structure of paraspeckles. Molecular cell. 2009; 33: 717-726.
39. Yang Z, Zhu Q, Luo K, Zhou Q. The 7SK small nuclear RNA inhibits the CDK9/cyclin T1 kinase to control transcription. Nature. 2001; 414: 317-322.
40. Markert A, Grimm M, Martinez J, Wiesner J, Meyerhans A, Meyuhas O, et al. The La-related protein LARP7 is a component of the 7SK ribonucleoprotein and affects transcription of cellular and viral polymerase II genes. EMBO reports. 2008; 9: 569-575.
41. Sedore SC, Byers SA, Biglione S, Price JP, Maury WJ, et al. Manipulation of P-TEFb control machinery by HIV: recruitment of P-TEFb from the large form by Tat and binding of HEXIM1 to TAR. Nucleic acids research. 2007; 35: 4347-4358.
42. Contreras X, Barboric M, Lenasi T, Peterlin BM. HMBA releases P-TEFb from HEXIM1 and 7SK snRNA via PI3K/Akt and activates HIV transcription. PLoS pathogens. 2007; 3: 1459-1469.
43. Willingham AT, Orth AP, Batalov S, Peters EC, Wen BG, et al. A strategy for probing the function of noncoding RNAs finds a repressor of NFAT. Science. 2005; 309: 1570-1573.
44. Sharma S, Findlay GM, Bandukwala HS, Oberdoerffer S, Baust B, et al. Dephosphorylation of the nuclear factor of activated T cells (NFAT) transcription factor is regulated by an RNA-protein scaffold complex. Proceedings of the National Academy of Sciences of the United States of America. 2011; 108: 11381-11386.
45. Li J, Chen C, Ma X, Geng G, Liu B, Zhang Y, Zhang S, et al. Long noncoding RNA NRON contributes to HIV-1 latency by specifically inducing tat protein degradation. Nature communications. 2016; 7: 11730.
46. Saayman S, Roberts TC, Morris KV, Weinberg MS. HIV Latency and the noncoding RNA therapeutic landscape. Adv Exp Med Biol. 2015; 848: 169 189.